Every day I sit on my porch and look at flowers, clouds and birds. I do this not once a day, but a lot. Early in the morning I drink a coffee and watch how the flowers open and the birds wake up, where the wind comes from and what the weather will be like.
During the day, I am watching everything around – how the flowers turn their heads towards the sun, how the sparrows in the feeder choose what is tastier, the magpies hatch the chicks, and the woodpecker seeks food in the old apple tree. The season of May bugs riot has just recently ended here. They are frantic in their desire to break into something. Continuous hum and constant blows on any surface. Now butterflies have appeared. For some reason, they really like the bush of white lilacs. Not tulips, not lilac lilacs, exactly white. And in the evening I watch the flowers close, moths crawl out, stars appear and bats fly out to hunt.
Suddenly, I was very interested in why some flowers close at night or in the rain, and some do not. Well, “why” is not so bad. But due to what. Moreover, it is believed that dandelions are closed before the rain. And I have seen many times that no. More precisely, sometimes yes, sometimes no. And the birds sometimes stop singing before the rain, and sometimes they don’t.
Obviously, the reason for this behavior of the flowers is a change in the environment – air temperature, humidity and other parameters. But they have no muscles – how do they do it? And how do their sensors work? And the most interesting is why not everyone is doing it.
I sat down to read.
Having studied the issue, I came to the conclusion that the reason why some flowers close at night and others not is in the history of the evolution of each particular species. I did not find any lists that can be analyzed.
The most common theories say that in this way plants protect their reproductive organs from cold / moisture / predators.
There are essentially two mechanisms. One is mostly used for the movement of flowers – when enough light gets in (in the morning or when the weather clears up, each plant has its own sufficiency criteria), the growth process of the inner part of the petal is activated more intensively than the outer and the flower opens. When the light intensity drops to a certain level, the outer part of the petal begins to grow faster than the inner one and the flower closes.
The second method is used by plants for the most part for leaves. It consists of a “muscle play” – some plants have special organs – part of the leaf or stem consists of cells that can grow in size and decrease. And in some plants, all cells have such properties. And so – with a change in environmental factors (humidity, temperature, mechanical stress, etc.), the “muscles”, for example, increase on the bottom of the sheet and the sheet folds. With the disappearance of external factors, the “muscles” on the outside of the leaf relax, and on the inside, they tighten and the leaf opens. The mechanisms for this in plants are different.
For me, the question was not fully clarified, but here we need to go chat with knowledgeable people.
If anyone is interested in more details about the mechanisms, then below I have collected information on the physiology of plant movement:
Various types of movements are inherent in plants, which are based on the functioning of contractile proteins, similar to what happens in animals, and only plant-specific movements based on increased stretching and changes in hydrostatic pressure in the cells.
Until the beginning of the 19th century, it was believed that the plant had neither sensitivity nor motor activity. J.B. Lamarck in 1909 in his work “Philosophy of Zoology” wrote that plants never in any of their parts have sensitivity, are unable to digest food and do not make movements under the influence of irritations. Indeed, plants have no nervous and muscular systems; plants, usually, are attached forms, but they all have the ability to move.
The movements of plants usually cannot be seen directly, since they occur very slowly. But with proper patience, you can notice that the tops of the shoots make circular motions (nutations), growing shoots and leaf blades turn in the direction of one-sided incident light (tropism), when the day and night change, flowers open and close (nastia). This can be observed using single-frame filming with showing at an accelerated pace. Some plants have rapid motor reactions (seismic), similar in speed to the movements of animals: slamming a leaf-trap in a flytrap, folding leaves when struck by a bashful mimosa, the movement of the stamens of a cornflower and barberry.
Nastias, or nastic movements (from anc.Greek: σπαθητός ‘compacted’) are movements of the dorsoventral organs of plants, which are caused by the characteristics of the plant itself and appear when exposed to environmental factors (temperature, light, humidity, etc.). Unlike tropisms, nastias are faster and occur in response to undirected, scattered environmental stimuli.
For example, saffron and tulip flowers open and close in response to a change in ambient temperature (thermonasty). In warmth, the growth of the inner side of the petals accelerates – and the flowers open, and in cold weather, the growth of their outer side accelerates – the flower closes.
The daily rhythms of opening and closing flowers and inflorescences are directly related to nastia. For example, goatsbeard’s baskets opened early in the morning usually close by 10–11 o’clock, and the flowers of a white water lily, on the contrary, are open only during the day. Corresponding infusions are caused by changes in temperature and humidity that occur during the day.
Nastias, not associated with tissue growth processes, are being caused by changes in the cells themselves. An example is the so-called “sleep of leaves” of oxalis; its triple leaves under diffused light are located in a horizontal plane, but when exposed to sunlight, they quickly close up to an “umbrella” (photonasty and thermonasty). The leaves of the bashful mimosa are folded with various kinds of tremors (seismonasty) and even with a light touch; while petioles of complex cirrus leaves of this plant wilt. Seismonasty not only protects the organs of the plant, but can also have important adaptive value: the opening of tobacco flowers in the evening is caused by the appearance of insects pollinating these plants, and in the insectivorous sundew the nastical movements of the leaf plate covered with mucus secreting glandular hairs occur upon landing of the insect on it’s plate, and after it has stuck to the mucus, the leaf edges quickly close, and the victim’s digestion process begins (this is how the sundew gets nitrogenous food).
Nastia is usually divided into positive and negative. In the mornings, under bright sunlight, dandelion’s inflorescences-baskets open, and when the illumination decreases, they close (positive photonasty). The flowers of fragrant tobacco open in the evening, with a decrease in illumination. This phenomenon is called negative photonasty.
Classification
For various types of nastic movements, specific terms are used:
Autonasty are spontaneous rhythmic movements of leaves that are not associated with changes in external conditions.
Niktinasty are plant movements associated with a combined change in both light and temperature. Such a combined effect occurs during day and night shifts. An example is leaf movement in some legume species.
Seismonasty – movements caused by touch, concussion, etc.
Thermonasty – movements that are caused by changes in temperature.
Turgor movements – are associated with a change in turgor. These include nictinastic leaf movements. So, the leaves of many plants are also characterized by rhythmic movements associated with a change in turgor in the cells of leaf pads.
Photonasty – movements that are caused by a change in illumination.
Question of reason:
Charles Darwin told that flowers that close at night protect themselves from freezing. Another theory says that niktinastia of the plants help them save energy and probably smell for daytime flowering. After all, it is during the day that insects pollinate them most of all. And at this time they are most active.
But there are scientists who believe that such a process does not allow pollen to accumulate morning dew and become heavy from this. Insects collect dry pollen more easily, and nikinastia contributes to the successful propagation of plants in nature. And one rather unusual theory claims that the process of niktinastia is a specially developed mechanism for protection from nocturnal predators. It is believed that flowers, closing their buds, create the illusion of grass and become invisible to nocturnal predators. After all, for example, owls kill a flower when they look for small animals in it.
Question of mechanism:
Existing methods of movement in plants can be classified as follows:
1) intracellular movements (movement of the cytoplasm and organelles);
2) locomotor movement of cells using flagella (taxis);
3) growth movements based on cell growth by extension (lengthening of axial organs-shoots and roots, leaf growth, circular nutations, tropisms, growth infusions);
4) apical growth (growth of pollen tubes, root hairs, moss protonema);
5) reversible turgor movements: stomata movements, nastias, seismonastys.
In this case, I am interested in the mechanism of growth movements and turgor movements.
Growth movements.
Stretch growth includes the formation of a central vacuole in the cell, the accumulation of osmotically active substances (ions, sugars, organic acids, etc.) in it, the absorption of water and the softening and stretching of the cell walls. Hydrostatic (turgor) pressure is the main force that stretches the cell wall. Stretching of the cell walls is supported by the inclusion of new polysaccharide molecules. The cell wall exerts a back pressure equal to turgor pressure on the contents of the cell, which determines the tone of the cell and tissue. Simultaneously with cell stretching, the synthesis of cytoplasmic components occurs – the number of mitochondria, ribosomes, and other intracellular structures increases. Regulation of stretching is carried out by the hormonal system. The main role in the regulation of growth by stretching is played by indolyl-3-acetic acid (IAA). Auxin is produced at the top of the shoot and, moving into the stretch zone, induces the growth of cells that are ready to stretch. In stretched cells, IAA interacts with receptors that are apparently localized in the plasmalemma, endoplasmic reticulum, and cytoplasm. One of the results of this interaction is the activation of the release of H+ ions from cells and the acidification of the space of the cell walls, while the hyperpolarization of the membrane potential develops. Thus, auxin activation of the H+ pomp of the plasmalemma is one of the earliest events in the induction of growth by stretching. By acidifying the space of cell walls, conditions are created for their loosening: displacement of Ca2 + from cell walls, weakening of part of hydrogen bonds, creation of pH (5.0 and lower), favorable for acid hydrolases activity in walls – wall polymer modifiers – all this increases the extensibility of the walls . At the same time, IAA in combination with the receptor reaches the nucleus and activates RNA synthesis in it. Activation of the synthesis of RNA and proteins is necessary to maintain the started growth by stretching. Energy costs for all these processes are provided by increased respiration.
If one or another environmental factor acts on the plant or its organ unidirectionally, then depending on the strength (concentration gradient of the substance, light, etc.) and the duration of the factor’s action, growth bending (tropism) appears either from the acting factor (negative) or in its direction (positive). Tropical bends occur due to changes in the transport of auxin and, possibly, other phytohormones. There are photo-, thermo-, electro-, trauma-, gravel-, hydro- and auto-tropisms. With the help of tropisms, such an orientation of organs in space is carried out that ensures the most efficient use of nutritional factors by the plant, and also serves to protect against harmful effects.
Phototropisms
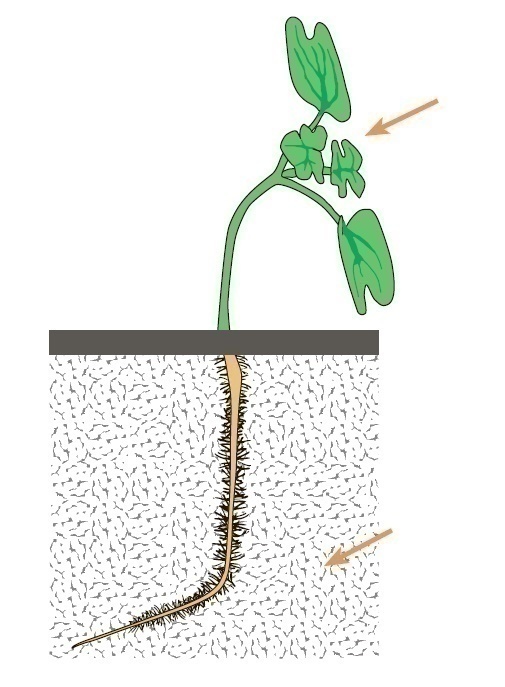
With positive phototropism, the green parts of the plant respond to the difference in the amount of light between the shadow and the illuminated sides of the organ (Fig. 1).
Fig. 1. Positive shoot phototropism and negative root phototropism in mustard seedling. The arrow indicates the direction of the light.
Negative phototropism is often observed at the roots, and the leaves may occupy a position perpendicular to the incident light. Phototropic bends are caused by the difference in the cell growth rate on two sides of the organ: the shaded side, usually, grows more intensively, since the polar transport of IAA is inhibited on the lit side, which moves to the shaded side. In addition, abscisic acid is synthesized on the lighted side, which, in turn, inhibits growth. The phototropic reaction is recorded in cereals after 20-25 minutes at a favorable temperature, and 1-3 hours after the start of one-sided illumination, both growth bending and transverse electrical polarization of tissues are manifested. The signal function in starting the phototropic reaction is performed by the light of the blue part of the spectrum (370, 425, 445, and 474 nm).
Reversible turgor movements.
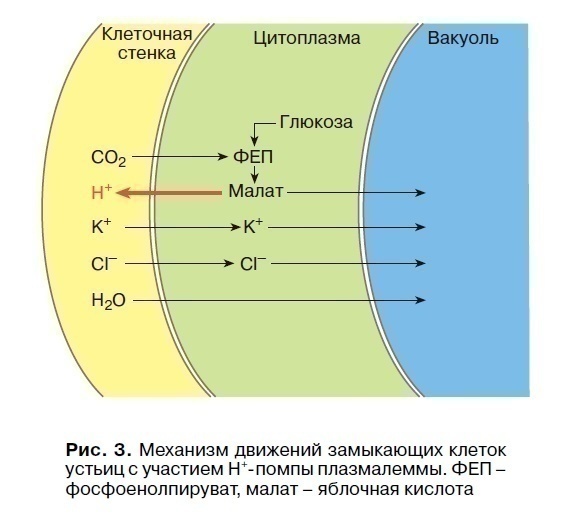
An example of slow turgor movements is the movement of trailing stomata cells that open or close the stomatal gap. The degree of opening of stomata depends on the illumination, the level of CO2 in the tissues of the leaf and their hydration. This is an important mechanism for regulating the gas and water regimes of the plant and the photosynthesis process carried out by the leaf. With sunrise, stomata opens at a low light intensity. They are especially sensitive to blue rays. Elucidation of molecular mechanisms showed that illumination of a plant adapted to darkness activates the operation of H+ pumps of the plasmalemma of stomata trailing cells (Fig. 3), as a result – their membrane potential is hyperpolarized. The exit of H+ ions from the trailing cell into the cell wall is accompanied by the simultaneous entry of K+ ions and Cl– anions from the walls into the cytoplasm and then into the vacuole. Anions of malic acid (malate) are transported to the vacuole from the cytoplasm. Following these ions, the flow of water into the vacuole increases, which increases the hydrostatic (turgor) pressure and the elastic walls of the trailing cells, stretching, open the stomatal gap. Phytohormones, auxins and cytokinins, activating the activity of H +, K + -ATPase plasmalemma, participate in the regulation of stomatal opening, and abscisic acid promotes the closing of stomata in conditions of insufficient hydration of leaf tissues.
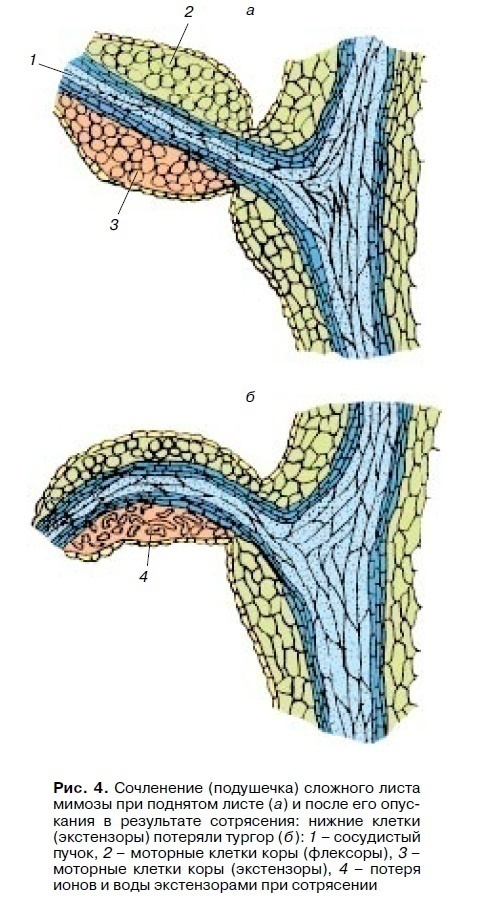
Similar mechanisms underlie all the turgor movements of plant organs in response to a change in diffusely acting environmental factors: light-darkness, heat-cold (nastia). An example is the movement of leaves of many legumes (beans, clover, peanuts, silk acacia) and some plants of other families (for example, oxalis) in response to a change of day and night. These movements are called nictinistic, or “sleep” movements. The movement of leaves in these plants is ensured by the activity of specialized formations – pads in the joints between the stem and petiole, between the petiole and leaf plate and between the leaves of a complex leaf (Fig. 4).
A vascular bundle passes through the center of the pillow, and around it (above and below) cortex cells are located, which serve as motor cells. When folded, some of them are able to lose turgor (extensors), while in others it increases (flexors). The upper and lower motor cells interact with each other. As in stomata, when the light regime changes, the activity of H + pumps changes, moving H+ ions out of motor cells and the transport of K+, Cl- and other ions begins. During movement, H+ pomp of motor cells of the upper and lower parts of the pads interact in opposite phases: with the onset of darkness, the activity of H+ pumps decreases in extensor cells, and their activity increases in flexor cells. Therefore, the nature of folding of leaves depends on the localization of extensors and flexors: if the extensor cells are located below the pads, then the leaves drop when shaded; if the extensors are localized at the top of the pads (like the leaves of a complex the bashful mimosa leaf), then the leaves will fold up.
The type of rapid turgor movements includes seismonastyc movements induced by concussion, impact of complex leaves of some legumes (mimosa), and venus flytrap. The most thoroughly studied seismonastyc response of bashful mimosa leaves. Mechanical shock (shaking) causes the appearance of an action potential (AP) in the leaf with an amplitude of 50-100 mV, which propagates through the petiole at a speed of 0.5-4 cm / s. At the point of transition of the petiole into the small pad, the initial AP induces a different action potential in the small pad. 0.05-0.1 s after the occurrence of AP in the pad, K + and Cl- begin to emerge, and after them water emerges from the extensor cells, which leads to a decrease in turgor in them. As a result, a complex leaf quickly drops (see Fig. 4), and small leaves rise and fold up (extensors from above). The whole leaf rises much more slowly (30-60 minutes). Such rapid motor reactions help prevent damage from strong winds and showers.
For seismonastyc movements, stages of absolute and relative immunity are also typical, during which, after a new stimulation, the excitation does not occur at all or appears only to a lesser extent. In the primary joint, the stage of absolute immunity lasts from 15 to 40 s, and the relative stage lasts about 360 s. During absolute immunity, Cl — K + exit the cell; at the same time, any subsequent equally strong irritation has no effect. During relative immunity, ion pumps begin to act again, and the Cl– and K + ions enter the cell again, but the action potentials cannot yet manifest in full force. Stages of immunity are being distinguished not only in relation to excitability, but also in relation to movements. This stage of absolute immunity continues until the response appears again (about 90 s). The stage of relative immunity (in relation to movements) ends only after 10-30 minutes, when the leaf returns to its original position. The recovery phase of the initial state is characterized by different steps. As a result of the implementation of processes that require energy, first of all, the semipermeability of membranes, the activity of ion pumps, and the initial ionic equilibrium are restored. Osmotic absorption of water occurs without energy consumption. It is especially interesting that the duration of the immunity stage does not depend on kind of involved irritation – mechanical, chemical, electrical, traumatic, or any other.
In addition to Mimosa pudica, many other plants can carry out seismonastyc movements of leaves. First of all, plants – inhabitants of the tropics. Representatives of the mimosa family Mimosa invisa and Neptunia oleracea, as well as the Oxalis family of Biophytum sensitivum and Oxalis sensitiva, can be mentioned as examples. Especially fast responses occur in Biophytum. They can be completed within 1s. The reverse movement lasts 3 minutes. Even strong concussions cause weak responses in these plants of our flora, and the “all or nothing” law does not apply to such movements. Between irritable and non-irritable objects, all transitional stages exist.
The evolution of modes of movement.
The question arises: how were such diverse types of movements formed in plants? It is generally accepted that higher plants evolved from green algae: Chlamydomonas, chlorococcal, ulotrix, chetoforov, first ground higher plants (rhinophytes), fern, gymnosperms and floral. Let us examine how the motor activity of plant organisms changed.
Monadic forms of green algae, living mainly in fresh waters, are moved with the help of flagella. Since flagella are cytoplasmic outgrowths, the cell walls of the monads are not completely closed and excess water is released from the cells using pulsating vacuoles. The cell membrane completely closes in chlorococcal algae, but at the same time they lose flagella and the ability to move independently. Multicellular filamentous algae (derived from chlorococcal) such as ulotrix are attached organisms, and their movement to the light falling from above is carried out slowly – due to cell division. The appearance of filamentous algae of a different, fast type of growth by the formation of a large central vacuole and extension of the cell wall — growth by extension, gave significant advantages in the mastering of better-lit upper layers of the reservoirs. Elongation of cells due to growth by stretching turned out to be such a successful form of movement that along with photosynthesis it became the basis for the development of the plant world. Other forms of movement in plants also include growth by stretching.
In the course of further evolution, the ability to reversible motions on the basis of changing turgor pressure develops increasingly in plants. Such a mechanism arose primarily in the first higher ground plants during the formation of stomata. Then slow turgor movements (nastia) begin to be used for leaf movement, closing and opening of flowers. Finally, fast reversible turgor movements appear – seismonastyc movements. If the regulation of movements associated with growth by stretching was carried out using phytohormones, then for the regulation of turgor movements the transmission rate of the hormonal signal is already insufficient, and the control of motor activity begins to be carried out with the participation of electrical impulses (action potentials). It is important to note that in plants of evolutionarily advanced taxa all the previously formed forms of movement are preserved in various combinations.
Hello! My name is Olga, nice to meet you!
In this blog, I am talking about our travels to beautiful and unusual places in Ukraine and beyond. Well and the history of these places. I sincerely believe that it is possible to find something interesting anywhere - just look at the outside world from different angles))) More
This website uses cookies to improve your experience. We'll assume you're ok with this, but you can opt-out if you wish. OKReject
Privacy & Cookies Policy
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are as essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.