Кожен день я сиджу у себе на ганку і дивлюся на квіти, хмари і птахів. Я це роблю не один раз в день, а багато. Рано вранці я п’ю кафе і дивлюся як відкриваються квіти і прокидаються птахи, звідки вітер і яка буде погода.
Вдень я спостерігаю за всім навколо – як квіти повертають свої голівки за сонцем, як горобці в годівниці вибирають що посмачніше, сороки висиджують пташенят, а дятел шукає прожиток в старій яблуні. Ось тільки недавно закінчився сезон буйства хрущів. Вони скажені в своєму прагненні розбитися об щось. Безперервний гул і постійні удари об будь-яку поверхню. Тепер з’явилися метелики. Чомусь їм дуже подобається кущ білого бузку. Не тюльпани, не звичайний бузок, а саме білий. А ввечері я дивлюся як квіти закриваються, виповзають метелики, з’являються зірки і вилітають на полювання кажани.
Раптом мене дуже зацікавило чому деякі квіти закриваються на ніч або під час дощу, а деякі – ні. Ну і “чому” це півбіди. А ось за рахунок чого. Причому, вважається, що кульбаби закриваються перед дощем. А я вже багато разів бачила, що немає. Точніше, іноді так, іноді ні. І птиці іноді перестають співати перед дощем, а іноді – ні.
Очевидно, що причина такої поведінки квітів в зміні навколишнього середовища – температури повітря, вологості та інших параметрів. Але у них же немає м’язів – як вони це роблять? І як у них влаштовані сенсори? А найцікавіше чому не всі цим займаються.
Сіла читати.
Вивчивши питання, я прийшла до висновку, що причина, з якої одні квіти закриваються на ніч, а інші ні – в історії еволюції кожного конкретного виду. Будь-яких списків, які можна проаналізувати, з цього приводу я не знайшла.
Найбільш поширені теорії говорять про те, що таким чином рослини захищають свої репродуктивні органи від холоду/вологи/хижаків.
Механізмів, по суті, два. Один здебільшого використовується для руху квітів – при попаданні достатньої кількості світла (вранці або проясненні погоди, у кожної рослини свої критерії достатності), активується процес зростання внутрішньої частину пелюстки інтенсивніше, ніж зовнішньої і квітка відкривається. При падінні інтенсивності освітлення до певного рівня, зовнішня частина пелюстки починає рости швидше внутрішньої і квітка закривається.
Другий спосіб використовується рослинами здебільшого для листя. Він полягає в “грі м’язами” – у деяких рослин є спеціальні органи – частина листа або стебла складається з клітин, здатних збільшуватися в розмірах і зменшуватися. А у деяких рослин такими властивостями володіють все клітини. І ось – при зміні факторів навколишнього середовища (вологість, температура, механічний вплив і ін.), “М’язи”, наприклад, на нижній частині листа збільшуються і лист складається. При зникненні зовнішніх факторів “м’язи” на зовнішній стороні листа розслабляються, а на внутрішній – напружуються і лист відкривається. Механізми для цього у рослин є різні.
Для мене питання прояснилося не до кінця, але тут треба піти поспілкуватися з досвідченими людьми.
Якщо кому цікаво докладніше про механізми, то нижче я зібрала інформацію з фізіології руху рослин:
Рослинам властиві різноманітні способи рухів, в основі яких лежать як функціонування скорочувальних білків, подібно до того, як це відбувається у тварин, так і властиві тільки рослинам руху на основі зростання розтягуванням і змін гідростатичного тиску в клітинах.
Аж до початку XIX століття вважалося, що рослина не володіє ні чутливістю, ні руховою активністю. Ж.Б. Ламарк в 1909 році в праці “Філософія зоології” писав, що рослини ніколи і в жодній зі своїх частин не володіють чутливістю, не мають здатності перетравлювати їжу і не роблять рухів під впливом подразнень. Дійсно, у рослин немає нервової і м’язової систем, рослини, як правило, є прикріпленими формами, але всі вони мають здатність до руху.
Рухи рослин зазвичай не вдається побачити безпосередньо, бо відбуваються вони дуже повільно. Але при належному терпінні можна помітити, що верхівки пагонів роблять кругові рухи (нутації), що ростуть пагони і листові пластинки повертаються в напрямку односторонньо падаючого світла (тропізми), при зміні дня і ночі відкриваються і закриваються квітки (настії). Це можна спостерігати за допомогою покадрової кінозйомки з показом в прискореному темпі. Деякі рослини володіють швидкими руховими реакціями (сейсмонастії), подібними по швидкості з рухами тварин: закриття листа-пастки у мухоловки, складання листів при ударі у мімози сором’язливої, рух тичинок волошки і барбарису.
Настії, або настічні рухи (від грец. Σπαθητός ‘ущільнений’) – рухи дорсовентральних органів рослин, які обумовлені особливостями самої рослини і проявляються при впливі факторів навколишнього середовища (температура, світло, вологість і ін.). На відміну від тропізмів, настії є більш швидкими і виникають у відповідь на ненаправлені, розсіяні в навколишньому середовищі подразники.
Наприклад, квітки шафрану і тюльпана відкриваються і закриваються у відповідь на зміну температури навколишнього середовища (термонастії). У теплі відбувається прискорення зростання внутрішньої сторони пелюсток – і квітки розкриваються, а при холоді відбувається прискорення зростання їх зовнішньої сторони – відбувається закриття квітки.
Безпосередньо пов’язані з настіями добові ритми відкривання і закривання квіток і суцвіть. Наприклад, відкриті рано вранці кошики козлобородника зазвичай закриваються до 10-11 години, а квітки білого латаття, навпаки, відкриті тільки вдень. Відповідні настії викликаються змінами температури і вологості протягом доби.
Настії, не пов’язані з процесами росту тканин, викликаються змінами самих клітин. Прикладом може служити так званий «сон листя» кислиці; її трійчасте листя на розсіяному світлі розташовується в горизонтальній площині, але при потраплянні сонячних променів вони швидко складаються «парасолькою» (фотонастії і термонастії). Листя ж мімози сором’язливої складаються при різного роду струсах (сейсмонастії) і навіть при легкому дотику; при цьому черешки складних перистих листів цієї рослини никнуть. Сейсмонастії не тільки забезпечують захист органів рослини, але можуть також мати важливе адаптивне значення: відкриття квіток тютюну у вечірній час обумовлено появою комах, що обпилюють дані рослини, а у комахоїдної росички настічні руху листової пластинки, покритої залозистими волосками, які виділяють слиз, виникають при посадці на цю пластинку комахи. Причому після того, як воно прилипло до слизу, края листа швидко закриваються, і починається процес перетравлення жертви (так росичка видобуває азотисту їжу).
Настії прийнято розділяти на позитивні і негативні. Вранці, при яскравому сонячному освітленні відкриваються суцвіття-кошики кульбаб, а при зменшенні освітленості відбувається їх закриття (позитивна фотонастія). Квітки запашного тютюну розкриваються у вечірній час, при зменшенні освітленості. Це явище називається негативною фотонастією.
Класифікація
Для різних типів настічних рухів використовуються особливі терміни:
Автонастії – мимовільні ритмічні рухи листя, не пов’язані зі змінами зовнішніх умов.
Ніктінастії – рухи рослин, пов’язані з комбінованими змінами, як освітленості, так і температури. Такий комбінований вплив настає при змінах дня і ночі. Прикладом є рух листя у деяких видів бобових.
Сейсмонастії – рухи, викликані дотиком, струсом і т.п
Термонастії – рухи, які викликані змінами температури.
Тургорні рухи – пов’язані зі зміною тургору. До них відносяться ніктінастічні рухи листя. Так, для листя багатьох рослин також характерні ритмічні рухи, пов’язані зі зміною тургору в клітинах листових подушечок.
Фотонастії – рухи, які викликані зміною освітленості.
Питання причини:
Чарльз Дарвін, стверджував, що квіти, які закриваються на ніч, оберігають себе від замерзання. Інша теорія говорить, що ніктінастія рослин допомагає зберегти їм енергію і ймовірно запах для денного цвітіння. Адже саме вдень найбільше літає комах, які запилюють їх. І в цей час вони найбільш активні.
А є вчені, які вважають, що такий процес не дає пилку набратися ранкової роси і ставати від цього важким. Комахи легше збирають сухий пилок, і ніктінастія сприяє успішному розмноженню рослин в природі. А одна досить незвичайна теорія стверджує, що процес ніктінастіі це спеціально розвинений механізм для захисту від нічних хижаків. Вважається, що квіти, закривши свої бутони, створюють ілюзію трави і стають малопомітними для нічних хижаків. Адже, наприклад, сови вбивають квітки, коли шукають в них дрібних тварин.
Питання механізму:
Існуючі способи руху у рослин можна класифікувати наступним чином:
1) внутрішньоклітинні рухи (рух цитоплазми і органоїдів);
2) локомоторні рухи клітин за допомогою джгутиків (таксиси);
3) ростові рухи на основі росту клітин розтягненням (подовження осьових органів-пагонів і коренів, ріст листя, кругові нутації, тропізми, ростові настії);
4) верхівкове зростання (зростання пилкових трубок, кореневих волосків, протонеми мохів);
5) оборотні тургорні рухи: рухи продихів, настії, сейсмонастії.
В даному випадку мене цікавить механізм ростових рухів і тургорного руху.
Ростові рухи.
Зростання розтягуванням включає в себе утворення в клітині центральної вакуолі, накопичення в ній осмотично активних речовин (іонів, цукрів, органічних кислот і ін.), поглинання води і розм’якшення і розтягнення клітинних стінок. Гідростатичний (тургорний) тиск є основною силою, що розтягує клітинну стінку. Розтягування клітинних стінок підтримується включенням в них нових молекул полісахаридів. Клітинна стінка надає на вміст клітини протитиск, рівний тургорному тиску, що визначає тонус клітини і тканини. Одночасно з розтягуванням клітини відбувається синтез компонентів цитоплазми – зростає число мітохондрій, рибосом та інших внутрішньоклітинних структур. Регуляція розтягування здійснюється гормональною системою. Основну роль в регуляції росту розтягуванням виконує індолил-3-оцтова кислота (ІОК). Ауксин виробляється в верхівці пагону і, переміщаючись в зону розтягування, індукує ріст клітин, готових перейти до розтягування. У клітинах що розтягуються, ІОК взаємодіє з рецепторами, які, мабуть, локалізовані в плазмалемі, ендоплазматичній мережі і цитоплазмі. Одним з результатів цієї взаємодії є активація виділення іонів Н + з клітин і підкислення простору клітинних стінок, одночасно розвивається гиперполяризація мембранного потенціалу. Таким чином, активація ауксином Н + -помпи плазмалеми – це одна з ранніх подій в індукції зростання розтягуванням. Підкисленням простору клітинних стінок створюються умови для їх розпушення: витіснення Са 2+ з клітинних стінок, ослаблення частини водневих зв’язків, створення рН (5,0 і нижче), сприятливого для діяльності в стінках кислих гідролаз – модифікаторів полімерів стінки, – все це збільшує розтяжність стінок . У той же час ІОК в комплексі з рецептором досягає ядра і активує в ньому синтез РНК. Активація синтезу РНК і білків необхідна для підтримки початку росту розтягуванням. Енергетичні витрати на всі ці процеси забезпечуються посиленням дихання.
Якщо той чи інший фактор середовища діє на рослину або його орган односпрямовано, то в залежності від сили (градієнта концентрації речовини, світла і т.ін.) і тривалості дії фактору виникає ростовий вигин (тропізм) або від діючого фактору (негативний), або в його сторону (позитивний). Тропічні вигини відбуваються внаслідок зміни транспорту ауксину і, можливо, інших фітогормонів. Розрізняють фото-, термо-, електро-, травмо-, граві-, гідро- і автотропізми. За допомогою тропізмов здійснюється така орієнтація органів в просторі, яка забезпечує найбільш ефективне використання рослиною факторів харчування, а також служить для захисту від шкідливих впливів.
фототропізм.
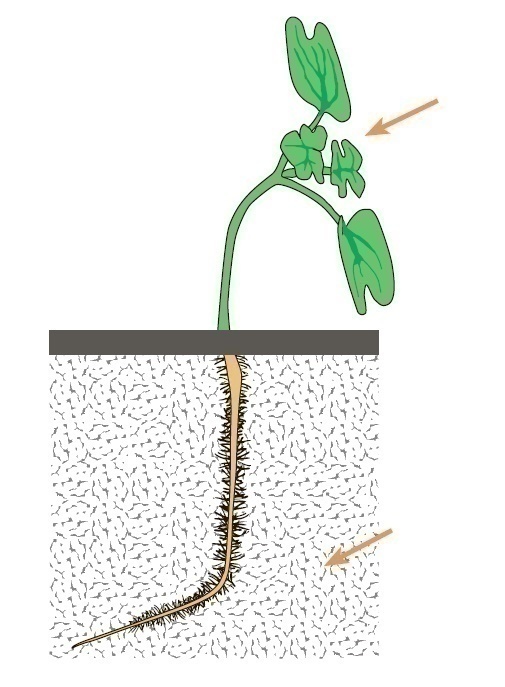
При позитивному фототропізм зелені частини рослини реагують на різницю в кількості світла між тіньовою і освітленою сторонами органу (мал. 1).
Мал. 1. Позитивний фототропізм пагону і негативний фототропізм кореня у проростка гірчиці. Стрілкою зазначений напрямок світла.
У коренів часто спостерігається негативний фототропізм, а листя можуть займати положення, перпендикулярне до падаючого світла. Фототропічні вигини обумовлені розходженням у швидкості росту клітин на двох сторонах органу: затінена сторона, здебільшого, зростає більш інтенсивно, тому що на освітленій стороні гальмується полярний транспорт ІОК, яка переміщається на затінену сторону. Крім того, на освітленій стороні синтезується абсцизова кислота, яка, в свою чергу, гальмує зростання. Фототропічна реакція реєструється у злаків через 20-25 хв при сприятливій температурі, а через 1-3 години після початку одностороннього висвітлення проявляється як ростовий вигин, так і поперечна електрична поляризація тканин. Сигнальну функцію в запуску фототропічної реакції виконує світло синьої частини спектра (370, 425, 445 і 474 нм).
Оборотні тургорні рухи.
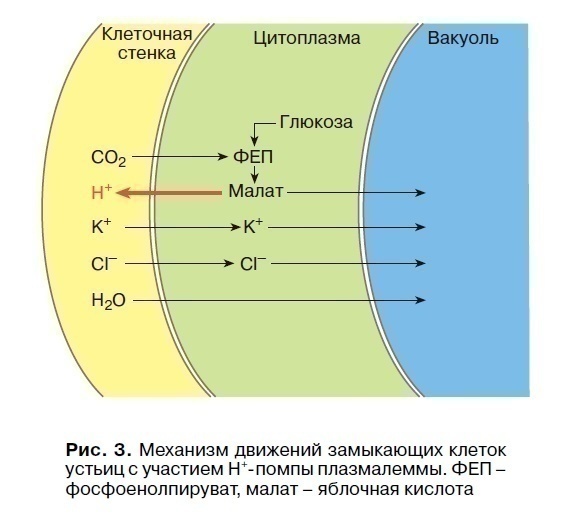
Прикладом повільних тургорних рухів можуть служити рухи замикаючі клітини продихів, які відкривають або закривають щілину продихів. Ступінь відкриття продихів залежить від освітленості, рівня СО2 в тканинах листа і їх зводненності. Це важливий механізм регуляції газового та водного режимів рослини і процесу фотосинтезу, здійснюваного листом. Зі сходом сонця продихи відкриваються при невеликій інтенсивності світла. Вони особливо чутливі до синіх променів. З’ясування молекулярних механізмів показало, що освітлення рослини, адаптованого до темряви, активує роботу Н + -насосів плазмалеми замикаючих клітин продихів, в результаті чого відбувається гиперполяризація їх мембранного потенціалу. Вихід іонів Н + з замикаючої клітини в клітинну стінку супроводжується одночасним входом іонів К + і аніонів Cl- зі стінок в цитоплазму і потім в вакуоль. Аніони яблучної кислоти (малати) транспортуються в вакуоль з цитоплазми. Слідом за цими іонами посилюється надходження води в вакуоль, що збільшує гідростатичний (тургорний) тиск і еластичні стінки замикаючих клітин, розтягуючись, відкривають щілину продиху. У регуляції відкривання продихів беруть участь фітогормони – ауксини і цитокініни, що активують діяльність Н +, К + -АТФази плазмалеми, а абсцизова кислота сприяє закриттю продихів в умовах недостатньої зводненості тканин листа.
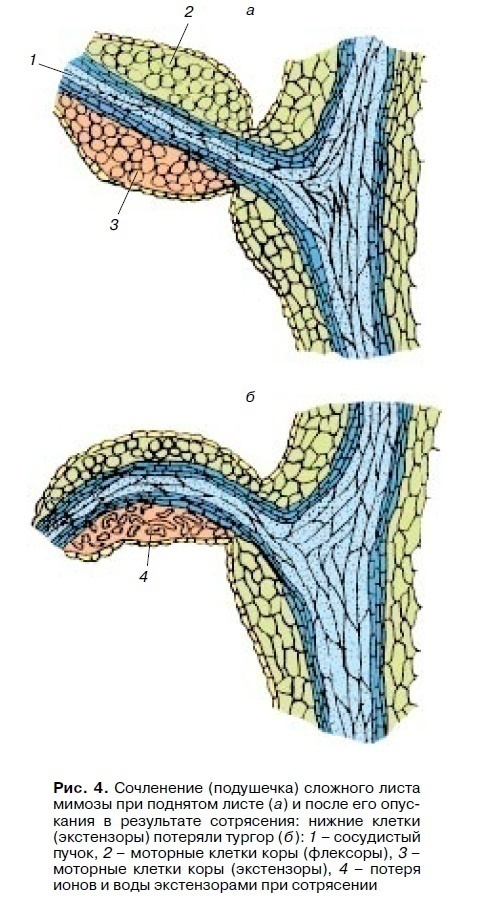
Подібні механізми лежать в основі всіх тургорних рухів органів рослин у відповідь на зміну дифузно діючих факторів зовнішнього середовища: світло-темрява, тепло-холод (настії). Прикладом можуть служити рухи листя багатьох бобових (квасолі, конюшини, арахісу, акації шовкової) і деяких рослин інших сімейств (наприклад, кислиці) у відповідь на зміну дня і ночі. Ці рухи називають ніктінастічними, або рухами “сну”. Рух листя у цих рослин забезпечується діяльністю спеціалізованих утворень – подушечок в зчленуваннях між стеблом і черешком, між черешком і платівкою листа і між листочками складного листа (мал. 4).
По центру подушечки проходить судинний пучок, а навколо нього зверху і знизу розташовуються клітини кори, які і виконують функцію моторних клітин. Одні з них при складанні листя здатні втрачати тургор (екстензори), тоді як в інших при цьому він зростає (флексори). Верхні і нижні моторні клітини взаємодіють між собою. Як і в продихах, при зміні режиму освітленості змінюється активність Н + -насосів, виводяться іони Н + з моторних клітин, і починається транспорт К +, Cl- та інших іонів. При русі взаємодіють Н + -помпи моторних клітин верхніх і нижніх частин подушечок, що працюють в протилежних фазах: з настанням темряви в екстензорних клітинах знижується активність Н + -насосів, а у флексорних клітин їх активність зростає. Тому від локалізації екстензорів і флексорів залежить характер складання листя: якщо екстензорні клітини знаходяться знизу подушечки, то листя при затіненні опускаються; якщо екстензори локалізовані вгорі подушечки (як у листочків складного листа мімози сором’язливої), то листя будуть складатися вгору.
До типу швидких тургорних рухів відносяться сейсмонастічні рухи, індуковані струсом, ударом складного листя деяких бобових (мімози), венериної мухоловки. Найбільш детально вивчена сейсмонастічна реакція листя мімози сором’язливої. Механічний удар (струшування) викликає в листі виникнення потенціалу дії (ПД) з амплітудою 50-100 мВ, який поширюється по черешку зі швидкістю 0,5-4 см/с. У місці переходу черешка в подушечку початковий ПД індукує в подушечці інший потенціал дії. Через 0,05-0,1 с після виникнення ПД в подушечці починається вихід К + і Cl- і слідом за ними води з екстензорних клітин, що призводить до зниження тургору в них. В результаті складний лист швидко опускається (див. мал. 4), а маленькі листочки піднімаються і складаються вгору (екстензори зверху). Весь лист піднімається значно повільніше (30-60 хв). Такі швидкі рухові реакції сприяють запобіганню пошкоджень від сильних вітрів і злив.
Для сейсмонастічних рухів типові також стадії абсолютної і відносної несприйнятливості, протягом яких після нового роздратування збудження зовсім не настає або проявляється лише в меншій мірі. У первинному зчленуванні стадія абсолютної несприйнятливості триває від 15 до 40 с, а відносна – приблизно 360 с. Під час абсолютної несприйнятливості Cl-К + виходять з клітини; при цьому будь-яке подальше настільки ж сильне роздратування не робить дії. Під час відносної несприйнятливості знову починають діяти іонні насоси, і іони Сl- і К + знову надходять в клітину, але потенціали дії ще не можуть проявитися в повну силу. Розрізняють стадії несприйнятливості не тільки по відношенню до збудливості, але і по відношенню до рухів. Така стадія абсолютної несприйнятливості триває до тих пір, поки знову не стане проявлятися відповідна реакція (близько 90 с). Стадія відносної несприйнятливості (щодо рухів) закінчується лише через 10-30 хв, коли лист повернеться в початкове положення. Для фази відновлення вихідного стану характерні різні ступені. В результаті здійснення процесів, що вимагають витрат енергії, в першу чергу відновлюються напівпроникливості мембран, діяльність іонних насосів і початкова іонна рівновага. Осмотичне ж поглинання води відбувається без витрат енергії. Особливо цікаво, що тривалість стадії несприйнятливості не залежить від того, про яке подразнення йдеться – механічне, хімічне, електричне, травматичне або будь-яке інше.
Крім Mimosa pudica, здійснювати сейсмонастічні рухи листя можуть і багато інших рослин. Перш за все це рослини – мешканці тропіків. Як приклади можна назвати представників сімейства мімоз Mimosa invisa і Neptunia oleracea, а також сімейства кісличних – Biophytum sensitivum і Охаlis sensitiva. Особливо швидко відповідні реакції відбуваються у Biophytum. Вони можуть завершитися протягом 1 с. Зворотний рух триває 3 хв. Навіть сильні струси викликають у цих рослин нашої флори слабкі відповідні реакції, і закон «все або нічого» до таких рухів непридатний. Між подразливими і не подразливими об’єктами існують всі перехідні ступені.
Еволюція способів руху.
Виникає питання: яким чином формувалися у рослин такі різноманітні типи рухів? Прийнято вважати, що вищі рослини походять від зелених водоростей: хламідомонадні, хлорококові, улотріксові, хетофорові, перші наземні вищі рослини (риніофіти), папоротеподібні, голонасінні і квіткові. Простежимо, як при цьому змінювалася рухова активність рослинних організмів.
Монадні форми зелених водоростей, що живуть в основному в прісних водах, пересуваються за допомогою джгутиків. Оскільки джгутики є цитоплазматичними виростами, то клітинні стінки у монад в повному обсязі замкнуті і надлишок води виділяється з клітин за допомогою пульсуючих вакуолей. Клітинна оболонка повністю замикається у хлорококових водоростей, але при цьому вони втрачають джгутики і здатність до самостійного пересування. Багатоклітинні нитчасті водорості (походять від хлорококових) типу улотрикса є прикріпленими організмами, і їх рух до падаючого зверху світла здійснюється повільно – за рахунок поділу клітин. Поява у нитчастих водоростей іншого, швидкого типу зростання шляхом утворення великої центральної вакуолі і розтягування клітинної стінки – зростання розтягуванням давало істотні переваги в освоєнні краще освітлених верхніх шарів водойми. Подовження клітин за рахунок зростання розтягуванням виявилося настільки вдалою формою руху, що поряд з фотосинтезом стало основою розвитку рослинного світу. Інші форми рухів у рослин також включають в себе зростання розтягуванням.
В ході подальшої еволюції у рослин все більшою мірою розвивається здатність до оборотних рухів на основі змінення тургорного тиску. Такий механізм виник перш за все у перших вищих наземних рослин при формуванні продихів. Потім повільні тургорні рухи (настії) починають використовуватися для руху листя, закривання і відкривання квіток. Нарешті, з’являються швидкі оборотні тургорні рухи – сейсмонастії. Якщо регуляція рухів, пов’язаних з ростом розтягуванням, здійснювалася за допомогою фітогормонів, то для регуляції тургорного руху швидкість передачі гормонального сигналу вже є недостатньою і управління руховою активністю починає здійснюватися за участю електричних імпульсів (потенціалів дії). Важливо відзначити, що у рослин еволюційно просунутих таксонів зберігаються в різних комбінаціях і всі раніше виниклі форми руху.
Добридень! Мене звуть Ольга, рада знайомству!
У цьому блозі я розповідаю про наші подорожі красивими і незвичайними місцями України і не тільки. Ну і історії цих місць. Я щиро вважаю, що у будь-якому місці можна знайти щось цікаве - для цього потрібно лише дивитися під різними кутами на навколишній світ))) Докладніше
Ми використовуємо кукі для найкращого представлення нашого сайту. Якщо Ви продовжите використовувати сайт, ми будемо вважати що Вас це влаштовує, але Ви можете відмовитися, якщо хочете.OKReject
Privacy & Cookies Policy
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these cookies, the cookies that are categorized as necessary are stored on your browser as they are as essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may have an effect on your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.